
Thực vật CAM là gì? Gồm các loài cây nào?
Thực vật CAM hay thực vật có cơ chế quang hợp kiểu CAM gồm những loài mọng nước, sống ở vùng hoang mạc khô hạn như: xương rồng, thanh long, dứa, sen đá... Bài viết dưới đây sẽ giúp bạn hiểu rõ hơn về thực vật CAM, cơ chế quang hợp của nó và các loại cây phổ biến thuộc loài này.
Thực vật CAM là gì?
Có những loài thực vật có cơ chế thích nghi đặc biệt để có thể phát triển tốt trong điều kiện khí hậu khô cằn như sa mạc, savan, núi đá (thực vật ưa khô hay thực vật chịu hạn). Một trong những cơ chế này gọi là cơ chế chuyển hóa axit Crassulacean viết tắt là CAM (Crassulacean acid metabolism) hay còn gọi là thực vật CAM.
Thực vật CAM đóng kín các khí khổng (lỗ thở) vào ban ngày nhằm giữ gìn nước không cho thoát ra ngoài. Vào ban đêm các khí khổng này sẽ được mở ra khi không khí lạnh và có nhiều độ ẩm hơn, cho phép chúng hấp thụ CO2 để sử dụng trong chuyển hóa cacbon thành chất hữu cơ (thường diển ra trong quá trình quang hợp).
 Sen đá được xếp vào loại thực vật CAM vì khả năng chịu hạn tốt cũng như cơ chế tích trự nước trong lá và thân
Sen đá được xếp vào loại thực vật CAM vì khả năng chịu hạn tốt cũng như cơ chế tích trự nước trong lá và thân
Một số đặc điểm nhận biết thực vật CAM
- Loài thực vật chịu hạn có các lá dày với tỷ số diện tích bề mặt nhỏ hơn so với thể tích.
- Chúng thường có lớp cutin dày để bảo vệ không bị khô héo trước ánh nắng mặt trời gay gắt.
- Các khí khổng (lỗ thở) có thể đóng và ban ngày hoặc bị chìm xuống thành các hốc lõm ngăn thoát hơi nước.
- Một số loại sẽ rụng lá vào mùa khô (không phải mùa lạnh).
- Thích hợp sống tại vùng có nhiệt độ cao (trên 30 độ) và ít CO2 (sa mạc, núi đá)
- Cây rất dễ bị thối rễ hoặc úng lá nếu tưới nhiều và đất không kịp thoát nước.
- Một số loại khác có thể lưu giữ nước trong các không bào (xương rồng, lan và dứa, sen đá).
Cơ chế quang hợp CAM
Thực vật CAM đóng kín các khí khổng trong thời gian ban ngày nhằm giữ gìn nước bằng cách ngăn cản quá trình thoát-bốc hơi nước. Các khí khổng sẽ được mở ra vào thời gian ban đêm lạnh và ẩm hơn, cho phép chúng hấp thụ CO2 để sử dụng trong quá trình cố định cacbon (chuyển hóa chất dinh dưỡng). Cơ chế CAM cho phép các loài thực vật này có thể phát triển bình thường trong các điều kiện môi trường mà nếu khác đi thì là quá khô hạn đối với sự phát triển của chúng, hay ít nhất ra là làm cho chúng có thể chịu đựng được các điều kiện cực kỳ khô hạn.
- Pha tối ở thực vật CAM gần giống với pha tối ở thực vật C4, điểm khác biệt là về thời gian:
- Ở thực vật C4, cả 2 chu trình của pha tối đều diễn ra vào ban ngày.
- Ở thực vật CAM thì chu trình đầu cố định CO2 tạm thời được thực hiện vào ban đêm khi khí khổng mở và chu trình Canvin tái cố định CO2 thực hiện vào ban ngày khi khí khổng đóng.
Cơ chế quang hợp CAM
Quá trình tổng hợp chất hữu cơ của thực vật CAM được bắt đầu khi hợp chất 3-cacbon là Photphoenolpyruvat được Cacboxylat hóa thành Oxaloaxetat và nó sau đó bị khử để tạo ra Malat. Thực vật CAM lưu trữ các trung gian 4-cacbon này cùng các hợp chất hữu cơ đơn giản khác trong các không bào của chúng. Muối malat dễ dàng bị phá vỡ thành Pyruvat và CO2, sau đó pyruvat được Photphorylat hóa để tái sinh Photphoenolpyruvat (PEP). Trong thời gian ban ngày, axít malic bị chuyển ra khỏi các không bào và bị phân tách ra để tạo thành CO2 sao cho nó có thể được enzym RuBisCO sử dụng trong chu trình Calvin-Benson trong chất nền đệm của lục lạp. Bằng cách này nó làm giảm tốc độ thoát-bốc hơi nước trong quá trình trao đổi khí.
Thực vật CAM có quá trình quang hợp gần giống với thực vật C4 nhưng năng suất lại thấp hơn rất nhiều vì vậy thực vật CAM thường phát triển khá chậm, nguyên nhân là vì:
- Điều kiện sống của chúng quá khắc nghiệt (nắng nóng, khô hạn, ít CO2), dẫn đến không thuận lợi cho quá trình quang hợp (tổng hợp chất dinh dưỡng).
- Do đặc điểm di truyền nên khả năng đồng hóa C02 của chúng kém hơn thực vật C4
- Pha sáng cần ánh sáng nhưng chúng thường đóng lỗ khí vào ban ngày nên tạo được ít ATP và NADPH, ảnh hưởng đến hiệu suất tổng hợp chất hữu cơ trong pha tối.
Ở một vài phương diện, CAM tương tự như kiểu quang hợp C4, ngoại trừ một điều là thực vật CAM không chứa các tế bào bó màng bọc. Thực vật C4 bắt CO2 trong một kiểu mô tế bào (thịt lá) và sau đó di chuyển nó tới kiểu mô khác (các tế bào bó màng bao) sao cho quá trình cố định cacbon có thể diễn ra thông qua chu trình Calvin-Benson. Ngoài ra, trao đổi chất C4 diễn ra liên tục khi còn có ánh nắng, trong khi CAM chỉ diễn ra hoàn toàn vào ban đêm.

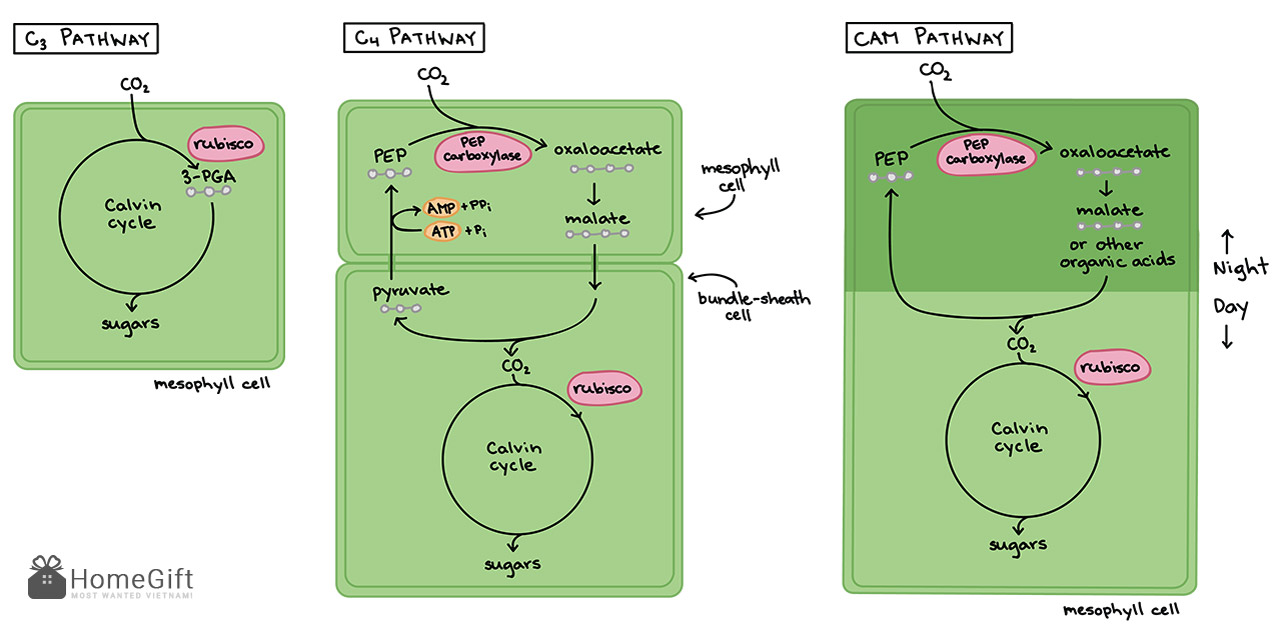
So sánh chu trình quang hợp của thực vật C3 C4 và CAM
Thực vật CAM có khả năng giữ nước rất tốt, cũng như rất hiệu quả trong việc sử dụng nitơ. Tuy nhiên, chúng là không hiệu quả trong việc hấp thụ CO2, do vậy chúng là các loại cây phát triển chậm khi so sánh với các loài thực vật khác. Ngoài ra, thực vật CAM cũng tránh quang hô hấp. Enzym chịu trách nhiệm cố định cacbon trong chu trình Calvin, Rubisco, không thể phân biệt CO2 với ôxy. Kết quả là thực vật sử dụng năng lượng để phá vỡ các hợp chất cacbon.
Các phân loại chính của thực Vật là gì?
Các loài thực vật trên hành tinh chúng ta rất đa dạng, tùy theo các điều kiện địa lý, khí hậu khác nhau chúng sẽ thích nghi thành nhiều loài khác nhau được phân thành 3 loại chính là thực vật C3, C4 và thực cật CAM. Dưới đây chúng ta sẽ tìm hiểu nhanh về 3 phân loại thực vật này nhé:
Sự khác nhau trong chu trình quang hợp giữa thực vật C3, C4 và CAM
Thực vật C3
Thực vật C3 là những thực vật mà sản phẩm ban đầu là 3-photphoglycerat với 3 nguyên tử cacbon. Những cây này còn được gọi là cây ôn đới. Những cây này khử thành khí cacbonic trực tiếp trong lục lạp.
Những cây này có thể sống lâu năm. Chúng rất giàu protein. Lúa mì, yến mạch, lúa mạch đen, cỏ vườn là một số ví dụ về thực vật C3.
Thực vật C3 gồm các loài rêu đến các cây gỗ lớn phân bố rộng khắp mọi nơi trên Trái Đất. Thực vật C3, có nguồn gốc từ đại Trung Sinh và đại Cổ Sinh, là xuất hiện trước thực vật C4 và hiện nay vẫn chiếm khoảng 95% sinh khối thực vật của Trái Đất. Thực vật C3 phải sinh sống tại các khu vực với nồng độ điôxít cacbon cao là do RuBisCO thông thường kết hợp phân tử ôxy vào RuBP thay vì phân tử điôxít cacbon.
Chu trình quang hợp Canvin ở thực vật C3
Theo Wiki: Cố định cacbon C3 là một kiểu trao đổi chất để cố định cacbon trong quang hợp ở thực vật. Quá trình này chuyển hóa điôxít cacbon và ribuloza bisphotphat (RuBP, một đường chứa 5-cacbon) thành 3-photphoglyxerat thông qua phản ứng sau:
6 CO2 + 6 RuBP → 12 3-photphoglyxerat
Thực vật C4
Thực vật C4 là thực vật chu trình cacbon đioxit thành các hợp chất đường 4 cacbon để đi vào chu trình C3 hoặc chu trình Calvin. Thực vật C4 có năng suất cao trong điều kiện khí hậu khô nóng và sản sinh nhiều năng lượng. Một số loại thực vật mà chúng ta thường tiêu thụ là thực vật C4 như dứa, ngô, mía, v.v.
Thực vật C4 bao gồm một số loại sống ở vùng nhiệt đới và cận nhiệt đới như Mía, Ngô, Cao lương (miến lúa). Thực vật C4 sống trong điều kiện nóng ẩm kéo dài, ánh sáng và nhiệt độ cao quanh năm nên các loài cây C4 có khả năng thích ứng nhiệt độ cao, cường độ quang hợp cao (cần nhiều ánh sáng), nhu cầu nước thấp (chịu hạn tốt).
Đặc điểm bên ngoài của dòng thực vật C4 là lá nhỏ và mảnh, chứa ít nước do vậy C4 ít bị mất nước và héo úa khi gặp nhiệt độ cao như các loại C3 (ngay cả khi bị cắt đứt ra khỏi thân thì là vẫn xanh trong nhiều giờ thậm chí nhiều ngày).
Chu trình quang hợp Hatch - Slack ở thực vật C4
Theo Wiki: Cố định cacbon C4 là một trong ba phương pháp, cùng với cố định cacbon C3 và quang hợp CAM, được thực vật trên đất liền sử dụng để "cố định" điôxít cacbon (liên kết các phân tử CO2 dạng khí thành các hợp chất hoà tan trong thực vật) để sản xuất đường thông qua quang hợp. Các loài thực vật sử dụng cơ chế cố định cacbon C4 được gọi chung là thực vật C4.
Thực vật CAM (Crassulacean acid metabolism)
Thực vật CAM là thực vật cố định carbon dioxide bằng con đường CAM hoặc chuyển hóa axit Crassulacean. Nó được phát hiện lần đầu tiên ở các loài thực vật thuộc họ Crassulaceae (họ lá bỏng). Chúng hiện diện trong môi trường khô và khô cằn. Con đường CAM được điều chỉnh để giảm thiểu thất thoát nước và phản xạ quang. Ví dụ về thực vật CAM bao gồm xương rồng, dứa, v.v.
Thực vật CAM chủ yếu là thực vật xerophytic (Cây chịu hạn). Con đường CAM cũng có trong một số loài thực vật thủy sinh như Hydrilla , Vallisneria,... Ở thực vật thủy sinh, con đường CAM xảy ra do sự khan hiếm CO2. Nguồn cung cấp carbon dioxide bị hạn chế do sự khuếch tán trong nước chậm hơn. Thực vật CAM thủy sinh hấp thụ CO2 vào ban đêm khi ít có sự cạnh tranh của các thực vật quang hợp khác.
Gồm những loài mọng nước, sống ở vùng hoang mạc khô hạn như: sen đá, xương rồng, dứa, thanh long… Tương tự thực vật C4 thì thực vật CAM cũng sống trong môi trường nhiệt độ cao, ánh sáng nhiều tuy nhiên độ ẩm thấp như sa mạc, savan, núi đá. Chính vì vậy cơ chế quang hợp của thực vật CAM rất đặc biệt. Để sống sót trong môi trường khắc nghiệt này, thực vật CAM phải có cơ chế tích trữ và chống mất nước. Trái với thực C4 là hạn chế nhu cầu nước thì thực vật CAM phải tích trữ nước trong lá và thân cây, thu hẹp diện tích tiếp xúc mặt trời bằng cách biến lá thành gai hoặc thu nhỏ, đặc điểm này làm cho nhiều loại thực vật CAM trông rất mọng và dày nên còn được gọi là loài mọng nước hoặc loại có gai như họ xương rồng.
Chu trình quang hợp (cố định Co2) gồm pha sáng và pha tối ở thực vật C3
Đặc điểm này cũng làm cho thực vật CAM trở thành loại cây phổ biến trong nghệ thuật terrarium (trồng cây trong lọ thủy tinh)
Thực vật CAM nhỏ xinh, mọng nước ít cần chăm sóc nên trở thành loại cây phổ biến trong nghệ thuật terrarium
Một số loại thực vật CAM phổ biến tại Việt Nam
Nguồn tham khảo:
- https://byjus.com/neet/short-notes-of-biology-for-neet-cam-plants/
- https://byjus.com/biology/difference-between-c3-and-c4-plants/
- https://biologydictionary.net/c3-c4-cam-plants/
- http://www.plantphysiol.org/content/155/1/56
- https://www.khanacademy.org/science/biology/photosynthesis-in-plants/photorespiration-c3-c4-cam-plants/a/c3-c4-and-cam-plants-agriculture
Link nội dung: https://vosc.edu.vn/nhung-cay-thuoc-nhom-thuc-vat-cam-la-a77057.html